

Research
Introduction:
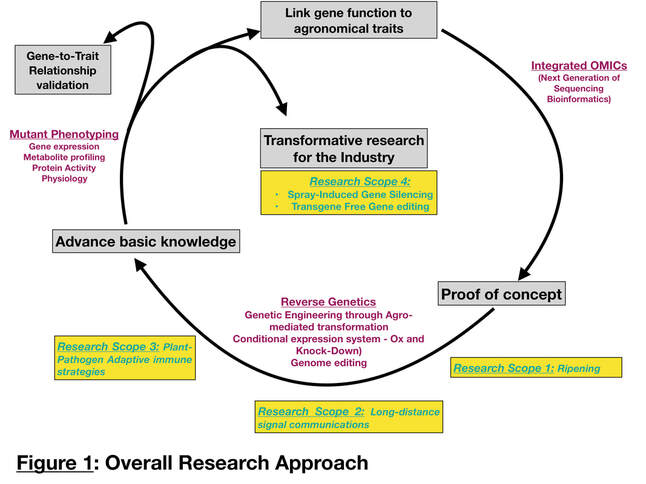
The overarching goal of the Deluc lab is to characterize the cause-to-effect relationships between genes and performance traits involved in plant development and responses to various stresses (Figure 1). Two models are mainly used in the lab; a perennial fruit crop (wine grape - Vitis Vinifera) and an annual crop (Brachypodium Distachyon). For both models, we have four main research scopes that are 1) Understanding the influence of auxin on the ripening process in grapevine, 2) Studying the role of signal molecules (hormone, peptides, microRNAs) in the scion/rootstocks interaction in the context of nutrient deficiencies and drought in grapevine and brachypodium, 3) Characterizing the role of co-(Alternative Splicing) and post-transcriptional events (microRNA) on the adaptive immunity of plants to confer tolerance to virus, and 4) Developing transformative research for crop protection (Spray Induced Gene Silencing and DNA-free CRISPR/Cas9 Genome Editing) in grapevine.
The overarching goal of the Deluc lab is to characterize the cause-to-effect relationships between genes and performance traits involved in plant development and responses to various stresses (Figure 1). Two models are mainly used in the lab; a perennial fruit crop (wine grape - Vitis Vinifera) and an annual crop (Brachypodium Distachyon). For both models, we have four main research scopes that are 1) Understanding the influence of auxin on the ripening process in grapevine, 2) Studying the role of signal molecules (hormone, peptides, microRNAs) in the scion/rootstocks interaction in the context of nutrient deficiencies and drought in grapevine and brachypodium, 3) Characterizing the role of co-(Alternative Splicing) and post-transcriptional events (microRNA) on the adaptive immunity of plants to confer tolerance to virus, and 4) Developing transformative research for crop protection (Spray Induced Gene Silencing and DNA-free CRISPR/Cas9 Genome Editing) in grapevine.
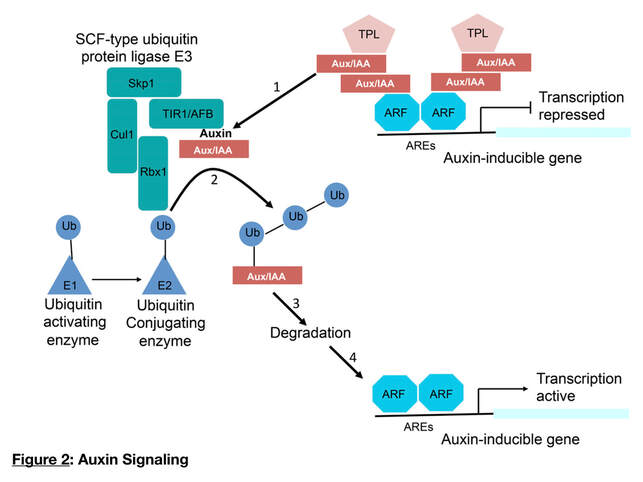
Research Scope 1 -The ripening process and auxin: Understanding the molecular mechanisms responsible for the genetic regulation of fleshy fruits is a major objective of fruit researchers. Fleshy fruits are divided into two main categories depending on whether the ripening process is controlled or not by the gaseous phytohormone "ethylene". Climacteric fruits such as tomato, apple, pear, and banana are characterized by a ripening-associated increase of respiration and the production of ethylene. By contrast, non-climacteric fruits, such as grape, orange, strawberry and pineapple lack this ethylene-associated respiratory peak. The signaling pathways that drive the ripening process in non-climacteric fruits remain elusive although the combined actions of ABA (Abscisic Acid), auxin, and sugar are suspected to be instrumental in the ripening initiation. In our lab, we use grape, as non-climacteric model, to characterize the influence of these different plant growth regulators with a particular focus on regulatory genes that trigger the major genetic "reprogramming" associated with the entry of grape berries to the ripening. Our scientific question is focused on the role of auxin during the ripening onset (Figure 2). The microvine (Chaib et al., 2010, Torregrosa et al., 2019), a grapevine model with a short life cycle, ever flowering, and easy to transform through Agrobacterium tumefaciens is used not only to validate the function candidate genes associated with Auxin, but also those that seems to interact with other regulators of plant growth regulators such as sugar, ABA, and brassinosteroids.
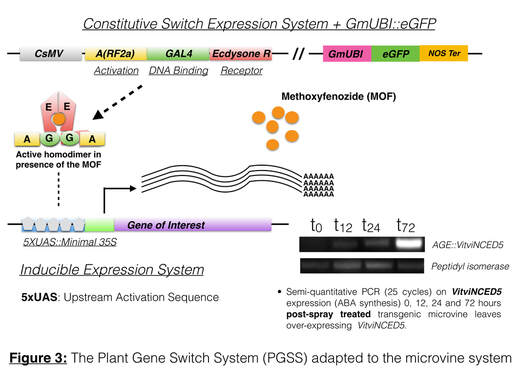
- Using this model system, we are currently validating the function of several genes involved in the signaling of Auxin (VitviARF4), and other ARF4 "interactors" associated with sugar, and ABA signaling. We conducted gain and loss-of-function studies on these different genes using a conditional expression system the Plant Gene Switch System (Martinez-Andujar et al., 2011 - Figure 3). This expression system enables us to decide when to induce the transgene (Ox and microRNA-driven Knock Down), which present some values when one wants understand the function of a gene in a temporal and spatial context.
- We also develop a research project focused on the influence of seed development the ripening. Previous works in the lab have demonstrated a relationship between seed mass relative to berry mass (Seed Index) with the timing of of ripening initiation (Gouthu and Deluc, 2015; Vondras et al., 2017). We have demonstrated an indirection relationship between the Seed Index and the entry of individual grape berries to the ripening regardless of the timing of anthesis (Vondras et al., 2016). High seed index berries delays the ripening while Low seed index berries are advanced. During the berry maturation phase, seeds experience a significant growth activity that is related to the differentiation of embryo. Production of plant growth regulators such as auxin is important in the seed during the berry maturation. We infer that a seed-driven auxin export to the pericarp explain the delay in berry with high seed weight. To answer this hypothesis, we have carried out two main projects aimed to understand the influence of seed on the ripening process. In the first research project, we carried out a genome-wide study to compare the transcriptional ripening programs of seed, pulp, and skin tissues of berries with high and low SI from berry maturation (pre-ripening phase) to berry maturity (harvest). Our research objective here is mainly oriented to gene discovery in order to identify what are the main regulatory "hubs" of the ripening program that are influenced by the seed development. The second project is to understand how seed development and embryo differentiation affects the movement and transport of auxin from the seed to the pericarp. For this, we are using the microvine as system to express fluorescent proteins (DR5:V2 and R2D2) that report auxin input and concentration in cells (Liao et al., 2015).
- The specific interplay between ABA, auxin and sugar is a critical component of the ripening initiation but the effects of genetic regulatory processes like epigenetic regulation are also explored. Some preliminary works in the lab indicate a potential interaction between Auxin signaling and a class of epigenetic factors (Demethylase: Jumonji Class). We are currently working in the development of transgenic lines using dCAs9 fused to either transcriptional activator or repressor under conditional expression (Lowder et al., 2015). Our objective is to spatially and temporally characterize three Jumonji genes at different transitional phases of grapevine growth (root, berries, shoot).
Research Scope 2 - Long-distance Signal communications between shoot and roots:
Long-distance signaling and communication between different organs of a plant is essential not only for the development and growth of the plant body, but also to adequately respond to unfavorable growing conditions. Higher plants coordinate and integrate the growth and development of their tissues and organs via sensory systems that results in the production of chemical, protein, nucleic, hydraulic, and electrical signals that are transported or spread from one part of the plant to another. In order to understand the impact of these molecules we adopt the technique of trans-grafting, which consists in disrupting a signal on one part of the plant (Genetically Engineered) and evaluate the impact on the other part (Non Genetically Engineered).
We are using two main models in the lab to study this topic:
Long-distance signaling and communication between different organs of a plant is essential not only for the development and growth of the plant body, but also to adequately respond to unfavorable growing conditions. Higher plants coordinate and integrate the growth and development of their tissues and organs via sensory systems that results in the production of chemical, protein, nucleic, hydraulic, and electrical signals that are transported or spread from one part of the plant to another. In order to understand the impact of these molecules we adopt the technique of trans-grafting, which consists in disrupting a signal on one part of the plant (Genetically Engineered) and evaluate the impact on the other part (Non Genetically Engineered).
We are using two main models in the lab to study this topic:
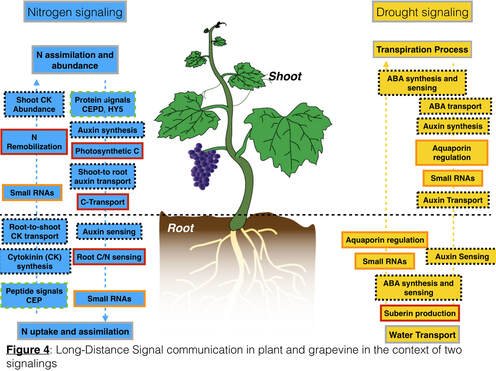
- The first one is microvine/grapevine model because of the extensively use of rootstock/scions in grapevine production. Several aspects of grapevine production including water stress, nutrient deficiency (Nitrogen, salt exclusion), and soil-borned disease relies on the use of rootstock and scion. However, genetic basis that confer drought and salt tolerance, excessive vigor, and disease resistance. Root-to-shoot and shoot-to-root communications are extremely relevant in such developmental context and in response to abiotic and biotic stresses (Figure 4). As first step, we have focused on two signals (chemical and hydraulic signals) in the context of drought.
- The second model is Brachypodium Distachyon. Our research question is oriented towards the influence of strigolactones, a plant hormone, on the control of root and shoot branching and the likely interplay of this class of regulators with other hormones under the context of nutrient deficiencies (Phosphate and Nitrate) and drought.
Research Scope 3 - Plant-Viral interactions:
Crop losses due to pest and diseases are a major threat to incomes rural families and to food security. The growing human population will require a significant increase in agricultural production from staple food to more marginal crops economically valuable though. Changes in the climactic and environmental conditions are leading to the appearance of new diseases whereas genetic changes within pathogen have resulted in the loss of effective sources of resistance and making the increasingly ineffective use of fungicides and pesticides a major concern for the sustainability of the environment of agricultural ecosystem. In this particular context, the identification of new resistance genes is mandatory. This will only be possible by a better understanding of the plant-pathogen interactions in order to produce either by conventional or molecular breeding new crop varieties that will carry a full or partial resistance without affecting other agronomically important traits. The interactions between a plant and its pathogen involve two-way communication by the ability of the plant to recognize and defense itself against a pathway and the capacity of the pathogen to circumvent the biology of the plant or to use it to its own benefit.
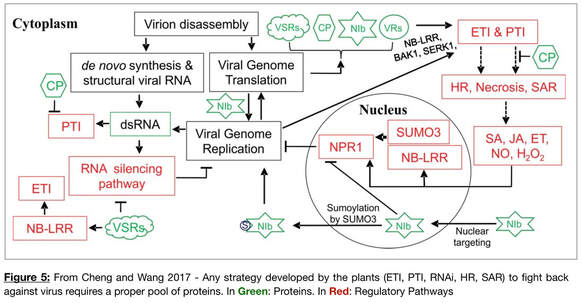
Among pathogens, plant viruses are extremely harmful to the production of vegetable, fruits, and cereal crops. Plant and viruses have developed various strategies (Figure 5). In the lab, we are studying the plant responses to two major viruses in grapevine (Grape Leafroll-associated Virus 3 (GLRaV3) and Grapevine Red Blotch Virus (GRBV). We are interesting in any aspect of transcriptional and post-transcriptional gene regulation, the impact of the viruses on plant alternatiev splicing and the type of host factors the viruses use to repliacet and move within the plants. The corresponding proteins are known to be co-opted by viral proteins to facilitate its spread. We believe that it is an additional strategy developed by the plant adaptive immune strategy to mitigate plant protein interactions with viral proteins, thereby limiting the spread of the virus.
Crop losses due to pest and diseases are a major threat to incomes rural families and to food security. The growing human population will require a significant increase in agricultural production from staple food to more marginal crops economically valuable though. Changes in the climactic and environmental conditions are leading to the appearance of new diseases whereas genetic changes within pathogen have resulted in the loss of effective sources of resistance and making the increasingly ineffective use of fungicides and pesticides a major concern for the sustainability of the environment of agricultural ecosystem. In this particular context, the identification of new resistance genes is mandatory. This will only be possible by a better understanding of the plant-pathogen interactions in order to produce either by conventional or molecular breeding new crop varieties that will carry a full or partial resistance without affecting other agronomically important traits. The interactions between a plant and its pathogen involve two-way communication by the ability of the plant to recognize and defense itself against a pathway and the capacity of the pathogen to circumvent the biology of the plant or to use it to its own benefit.
Among pathogens, plant viruses are extremely harmful to the production of vegetable, fruits, and cereal crops. Plant and viruses have developed various strategies (Figure 5). In the lab, we are studying the plant responses to two major viruses in grapevine (Grape Leafroll-associated Virus 3 (GLRaV3) and Grapevine Red Blotch Virus (GRBV). We are interesting in any aspect of transcriptional and post-transcriptional gene regulation, the impact of the viruses on plant alternatiev splicing and the type of host factors the viruses use to repliacet and move within the plants. The corresponding proteins are known to be co-opted by viral proteins to facilitate its spread. We believe that it is an additional strategy developed by the plant adaptive immune strategy to mitigate plant protein interactions with viral proteins, thereby limiting the spread of the virus.
Research Scope 4 - Transformative research:
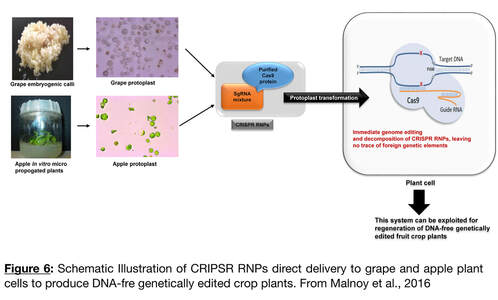
- Developing a Marker-Free Genome Editing methodology for clonally-propagated crop:
- Improving the generation of grapevine mutants to accelerate gene-to-trait relationship:
- Developing A Spray Induced Gene Silencing methodology for crop protection and improved fruit composition:
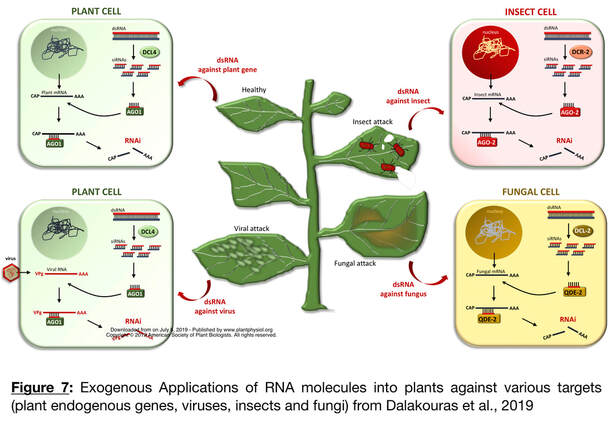
dsRNA molecules sprayed on the leaf surface were shown to be taken up by plant cells, transported to non-treated areas, and processed into sRNAs. These findings indicate that these applications can have systemic activity further enhancing their utility for pest and disease control. The premise is thus to deliver dsRNAs with a formulation made with clay particles using the same spray technology used for fungicides. Once applied, the dsRNA molecules complementary to target genes will disrupt the expression of the dsRNA-targeted genes in either the plant itself or the pathogens. The efficacy of SIGS was demonstrated for pest protection in multiple crops such as wheat, cucumber, pepper, etc.; and large-scale field trials are currently evaluated in Australia. The dsRNAs are reported to be stable and afford protection against the pathogen for up to 7-10 days, after which they degrade and leave no residue. The Fungus Erysiphe necator, responsible for Grape Powdery Mildew (GPM), is a significant threat to wine production that is mainly controlled using large quantities of fungicides. However, the emergence of fungicide resistance to chemicals suggests the need for rapid development of new strategies to control GPM. There is also a concern about the environmental impact of large-scale fungicide applications. Recent research demonstrates that agricultural pests, such as insects, nematodes, and fungi can be killed by exogenously supplied RNA molecules (Figure 7) opens opportunities to develop alternative field applications to control Powdery Mildew. We developed a SIGS to control GPM, we propose to accomplish three objectives.
In the first objective, we identified the stretches of RNA sequences with the maximum interference activity to silence fungal and native genes, EnCYP51, EnDCL1,2, and VitviMLOs 6,7, 11, respectively. In the second objective, we are currently evaluating the uptake and processing of RNA molecules by the grapevine and their systemic effects on the reduction of GPM. Finally, we will determine the efficacy of clay nanoparticles to stabilize and extend the lifespan of sprayed dsRNA molecules on grapevine leaves. Overall, we hope to provide the grape community with an RNAi-based spray technique for disease control. Our study will evaluate various aspects including the efficacy of silencing on the target genes, one adjuvant to use for long-lasting effects of dsRNA applications, the level of systemic effects, and the degree of PM-resistance afforded by the system in the laboratory studies. The results will be used for the development of field trials to evaluate the impact of SIGS system in reducing the cost of grapevine GPM management.
Citations:
Chaïb J, Torregrosa L, Mackenzie D, Corena P, Bouquet A, Thomas MR (2010) The grape microvine - a model system for rapid forward and reverse genetics of grapevines. Plant J 62: 1083–1092
Costa LD, Malnoy M, Lecourieux D, Deluc L, Lecourieux FO-, Thomas MR, Torregrosa LJ-M (2019) The state-of-the-art of grapevine biotechnology and new breeding technologies (NBTS): 1 53: 189–212
Dalakouras A, Wassenegger M, Dadami E, Ganopoulos I, Pappas M, Papadopoulou KK (2020) GMO-free RNAi: exogenous application of RNA molecules in plants. Plant Physiology pp.00570.2019
Gouthu S, Deluc LG (2015) Timing of ripening initiation in grape berries and its relationship to seed content and pericarp auxin levels. BMC Plant Biol 15: 46
Liao C-Y, Smet W, Brunoud G, Yoshida S, Vernoux T, Weijers D (2015) Reporters for sensitive and quantitative measurement of auxin response. Nature Methods 12: 207–210
Lowder LG, Zhang D, Baltes NJ, Paul JW, Tang X, Zheng X, Voytas DF, Hsieh T-F, Zhang Y, Qi Y (2015) A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol 169: 971–985
Malnoy M, Viola R, Jung M-H, Koo O-J, Kim S, Kim J-S, Velasco R, Nagamangala Kanchiswamy C (2016) DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front Plant Sci. doi: 10.3389/fpls.2016.01904
Numata K, Horii Y, Oikawa K, Miyagi Y, Demura T, Ohtani M (2018) Library screening of cell-penetrating peptide for BY-2 cells, leaves of Arabidopsis, tobacco, tomato, poplar, and rice callus. Scientific Reports 8: 10966
Ramakrishna S, Kwaku Dad A-B, Beloor J, Gopalappa R, Lee S-K, Kim H (2014) Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Research 24: 1020–1027
Vondras AM, Gouthu S, Schmidt JA, Petersen A-R, Deluc LG (2016) The contribution of flowering time and seed content to uneven ripening initiation among fruits within Vitis vinifera L. cv. Pinot noir clusters. Planta 243: 1191–1202
Chaïb J, Torregrosa L, Mackenzie D, Corena P, Bouquet A, Thomas MR (2010) The grape microvine - a model system for rapid forward and reverse genetics of grapevines. Plant J 62: 1083–1092
Costa LD, Malnoy M, Lecourieux D, Deluc L, Lecourieux FO-, Thomas MR, Torregrosa LJ-M (2019) The state-of-the-art of grapevine biotechnology and new breeding technologies (NBTS): 1 53: 189–212
Dalakouras A, Wassenegger M, Dadami E, Ganopoulos I, Pappas M, Papadopoulou KK (2020) GMO-free RNAi: exogenous application of RNA molecules in plants. Plant Physiology pp.00570.2019
Gouthu S, Deluc LG (2015) Timing of ripening initiation in grape berries and its relationship to seed content and pericarp auxin levels. BMC Plant Biol 15: 46
Liao C-Y, Smet W, Brunoud G, Yoshida S, Vernoux T, Weijers D (2015) Reporters for sensitive and quantitative measurement of auxin response. Nature Methods 12: 207–210
Lowder LG, Zhang D, Baltes NJ, Paul JW, Tang X, Zheng X, Voytas DF, Hsieh T-F, Zhang Y, Qi Y (2015) A CRISPR/Cas9 Toolbox for Multiplexed Plant Genome Editing and Transcriptional Regulation. Plant Physiol 169: 971–985
Malnoy M, Viola R, Jung M-H, Koo O-J, Kim S, Kim J-S, Velasco R, Nagamangala Kanchiswamy C (2016) DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front Plant Sci. doi: 10.3389/fpls.2016.01904
Numata K, Horii Y, Oikawa K, Miyagi Y, Demura T, Ohtani M (2018) Library screening of cell-penetrating peptide for BY-2 cells, leaves of Arabidopsis, tobacco, tomato, poplar, and rice callus. Scientific Reports 8: 10966
Ramakrishna S, Kwaku Dad A-B, Beloor J, Gopalappa R, Lee S-K, Kim H (2014) Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Research 24: 1020–1027
Vondras AM, Gouthu S, Schmidt JA, Petersen A-R, Deluc LG (2016) The contribution of flowering time and seed content to uneven ripening initiation among fruits within Vitis vinifera L. cv. Pinot noir clusters. Planta 243: 1191–1202